- Database
- Open access
- Published:
The Gene Ontology (GO) Cellular Component Ontology: integration with SAO (Subcellular Anatomy Ontology) and other recent developments
Journal of Biomedical Semantics volume 4, Article number: 20 (2013)
Abstract
Background
The Gene Ontology (GO) (http://www.geneontology.org/) contains a set of terms for describing the activity and actions of gene products across all kingdoms of life. Each of these activities is executed in a location within a cell or in the vicinity of a cell. In order to capture this context, the GO includes a sub-ontology called the Cellular Component (CC) ontology (GO-CCO). The primary use of this ontology is for GO annotation, but it has also been used for phenotype annotation, and for the annotation of images. Another ontology with similar scope to the GO-CCO is the Subcellular Anatomy Ontology (SAO), part of the Neuroscience Information Framework Standard (NIFSTD) suite of ontologies. The SAO also covers cell components, but in the domain of neuroscience.
Description
Recently, the GO-CCO was enriched in content and links to the Biological Process and Molecular Function branches of GO as well as to other ontologies. This was achieved in several ways. We carried out an amalgamation of SAO terms with GO-CCO ones; as a result, nearly 100 new neuroscience-related terms were added to the GO. The GO-CCO also contains relationships to GO Biological Process and Molecular Function terms, as well as connecting to external ontologies such as the Cell Ontology (CL). Terms representing protein complexes in the Protein Ontology (PRO) reference GO-CCO terms for their species-generic counterparts. GO-CCO terms can also be used to search a variety of databases.
Conclusions
In this publication we provide an overview of the GO-CCO, its overall design, and some recent extensions that make use of additional spatial information. One of the most recent developments of the GO-CCO was the merging in of the SAO, resulting in a single unified ontology designed to serve the needs of GO annotators as well as the specific needs of the neuroscience community.
Background
The Gene Ontology (GO) [1, 2] contains a set of terms for describing the activity and actions of gene products across all kingdoms of life. Each of these activities is executed in a cellular location or a location outside in the vicinity of a cell. In order to capture this context, the GO includes, since its inception, a sub-ontology called the Cellular Component Ontology (GO-CCO). GO-CCO terms describe parts of cells and structures associated with cells throughout the taxonomy range. The primary use of this ontology is for GO annotation, but it has also been used for phenotype annotation. Another ontology with a similar scope to the GO-CCO is the Subcellular Anatomy Ontology (SAO) [3], part of the Neuroscience Information Framework Standard (NIFSTD) [4] suite of ontologies. The SAO covers cellular components in the domain of neuroscience and was designed as a model for describing relationships among subcellular structures that would be encountered in an electron micrograph, for example a neuropil. In the nervous system, there are numerous examples of named subcellular structures that are composed of parts of multiple cell types, e.g., synapses, the Node of Ranvier, the glia limitans. SAO thus has a richer set of spatial relationships than the GO, modeled in part after the Foundational Model of Anatomy (FMA) [5].
At the time the SAO was constructed, circa 2005–2006, tools for import and reuse of existing ontologies were limited; in addition the SAO was composed in OWL (Web Ontology Language), while the GO-CCO was in OBO (Open Biomedical Ontologies) format. At this time, the semantics of OBO format were not yet aligned with those of OWL. Thus, the SAO had developed an independent set of cell component terms, with a heavy focus on those encountered in the nervous system. More recently, with the advent of a more detailed specification of OBO format (which clarifies the semantics of OBO format as a subset of OWL2) and the development of OBO/OWL converters, the native format of an ontology is less relevant. This has allowed us to work together on the same ontology by incorporating the SAO into the GO-CCO.
The SAO was used primarily within prototype segmentation and annotation tools developed for electron tomography data [6] to enhance search within the NIF across federated data [4] and, as described below, to annotate data derived from imaging and the literature on phenotypes associated with neurodegenerative disease [7]. To ensure that these annotations are not lost, NIF maintains a mapping between SAO and GO-CCO within a bridge file (for details on the use of bridge files in NIFSTD, see [8]).
In this paper, we describe an overview of the GO-CCO, a description of the amalgamation of the GO-CCO with the SAO, followed by a sketch of how the GO-CCO fits in with other ontologies. The last part of the paper describes applications and uses of the GO-CCO. Our aim is to provide a single unified cellular component ontology that can serve the needs of a diverse scientific community. The biomedical and bioinformatics communities may also benefit from the links between the GO-CCO and other ontologies.
The URL for the Gene Ontology (GO) is http://www.geneontology.org/. GO files are publicly available for download at http://geneontology.org/GO.downloads.ontology.shtml.
Overview of the Cellular Component Ontology
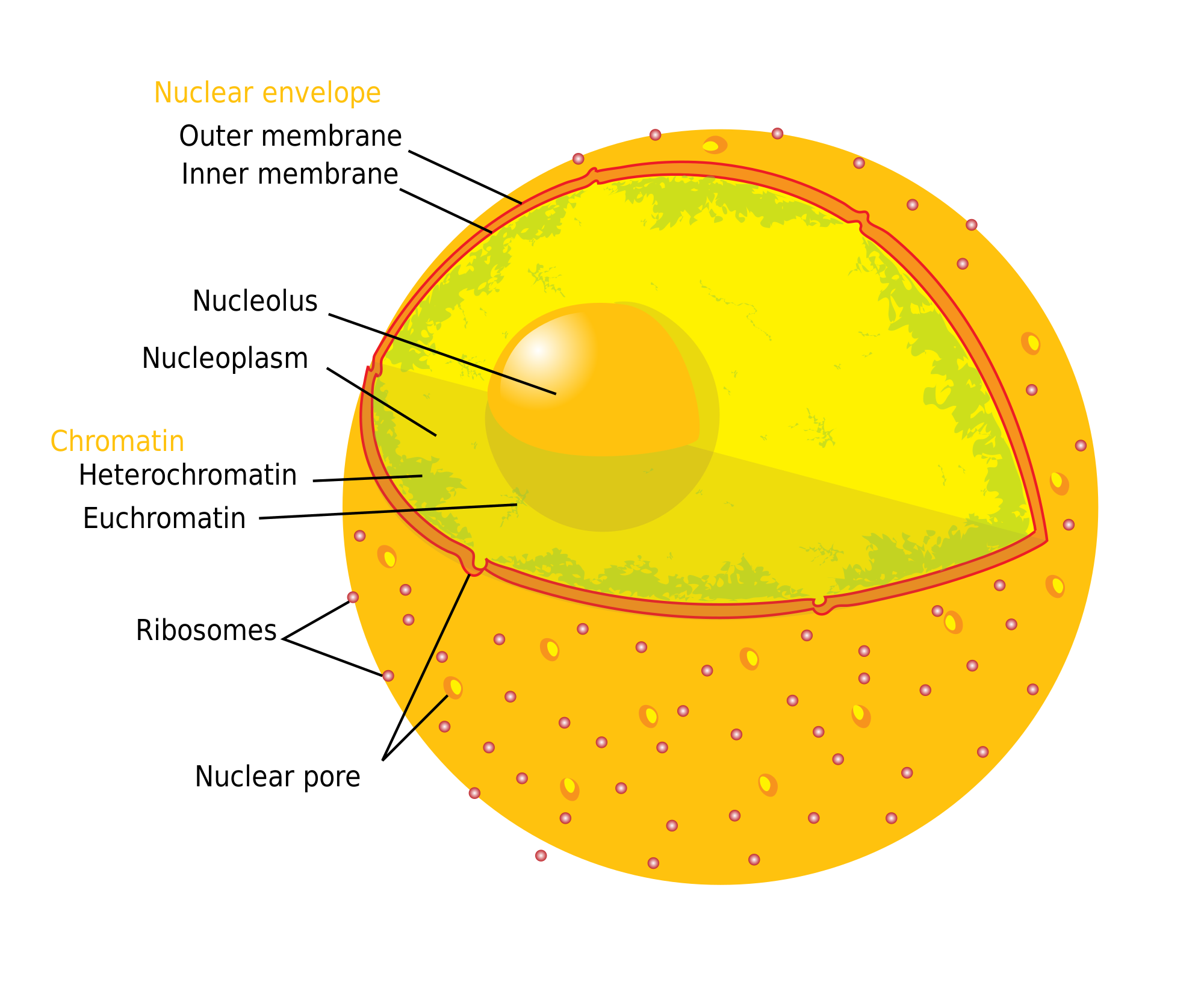
The Cellular Component Ontology describes subcellular structures and macromolecular complexes. GO-CCO terms may thus be used to annotate cellular locations of gene products. Examples of cellular components include ‘nuclear inner membrane’ (Figure 1) and the ‘ubiquitin ligase complex’, with several subtypes of this complex represented as descendants. The GO-CCO is not taxonomically restricted, and includes terms for both core components found across all domains of life (for example, the species-generic ‘chromosome’) and components specific to particular lineages (for example, ‘Nebenkern’, a mitochondrial formation found in insects, and ‘thylakoid’, a compartment inside chloroplasts and cyanobacteria).

Diagram and ontology placement of 'nuclear inner membrane'. (A) Diagram of human cell nucleus, including the nuclear inner membrane. (Taken from Wikimedia commons, http://upload.wikimedia.org/wikipedia/commons/thumb/3/38/Diagram_human_cell_nucleus.svg/2000px-Diagram_human_cell_nucleus.svg.png). (B) Placement of the Gene Ontology term GO:0005637 'nuclear inner membrane', drawn using the ontology editing tool OBO-Edit (see ‘Methods’). Due to space limitations, not all ancestor and descendant terms are shown. Is_a links are indicated by "I"; part_of links are indicated by "P" (see main text for explanation).
{kind=link}
The two core relationship types used in the GO-CCO are ‘is_a’ and ‘part_of’. The ‘is_a’ relation (also known as “SubClassOf”) represents the relationship between a more generic term and a specialized term (for example, between ‘membrane’ and ‘plasma membrane’), whereas the ‘part_of’ relationship describes how sub-structures are assembled into larger structures (for example, between ‘nucleolus’ and ‘nucleus’) [9].
Generally, experimental results or computational predictions support statements that a gene product is located in or is a subcomponent of a particular cellular component. The GO-CCO includes multi-subunit enzymes and other protein complexes, but not individual proteins or nucleic acids. (Terms describing protein complexes are further discussed below.) Whilst the GO-CCO includes cell structures, it excludes cell types, which are instead represented in the Cell Ontology (CL) [10] or the plant cell branch of the Plant Ontology (PO) [11]. The GO-CCO also excludes multicellular anatomical terms, with such structures being described by either species-specific ontologies (e.g., Zebrafish anatomy ontology [12], Mouse gross anatomy ontology [13]) or taxonomically broad anatomical ontologies (e.g., Uberon [14], PO).
The 2013-06-18 release of the GO contains 3332 CC ontology terms. Approximately half of these terms represent protein complexes, with the other half representing larger units.
Amalgamation with SAO
The SAO was incorporated into the Neuroscience Information Framework standard ontologies when they were originally assembled (NIFSTD) [15]. The NIF project [16] was charged with providing a semantic framework for describing and searching neuroscience data. NIFSTD was built from community ontologies when possible, but as noted above, working with community ontologies was often a challenge when the project began. Over the course of the project, NIF gradually replaced its custom ontologies with more general community ontologies when they became available, both to benefit from the continued enrichment of these ontologies by the life sciences community and to ensure that annotations in the NIF would be compatible with the larger life sciences community. In this case, a reconciliation of the NIF and the GO-CCO was required. Through this reconciliation, not only would NIF’s data federation and search benefit from the on-going development and extensive use of the GO for annotations, but the community ontologies would become enriched with the neuroscience-specific content developed by NIF. The SAO-GO-CCO integration is an example of this type of harmonization.
We started from a list of about 400 terms from the NIF Subcellular Anatomy Ontology (SAO) representing sub-cellular locations that required integration into the GO-CCO. GO editors carefully examined the list and considered each term as appropriate. The following categories were identified:
-
1)
Terms that were already in the GO;
-
2)
Terms that needed to be added to the GO;
-
3)
Terms that were out of scope for the GO.
Terms that were already in the GO were:
-
high-level GO-CCO terms that were included in the SAO to provide some structure (e.g. ‘plasma membrane’);
-
recent additions to the GO that had not yet been documented in SAO; in these cases, the NIFSTD IDs have been included in the GO as database cross-references;
-
present in the GO under a different primary name than used by the SAO/NIF; where appropriate, the missing SAO names have been added to the GO as synonyms, along with their NIFSTD IDs.
SAO terms that needed to be added to the GO were created, and their NIFSTD IDs recorded as database cross-references. Definitions for the SAO terms were taken from the SAO where available, edited if necessary, or written by GO editors based on the literature or after consultation with SAO editors. 97 new terms were added to the GO (e.g. ‘dendritic tree’ (Figure 2), ‘ribbon synapse’); their full list is shown in Table 1.

Image and ontology placement of ‘dendritic tree’. (A) Dendritic tree from a Purkinje neuron from mouse cerebellum injected with Lucifer Yellow and imaged using confocal microscopy. (Taken from the Cell Centered Database (CCDB), http://ccdb.ucsd.edu/sand/main?mpid=3&event=displayRecon). (B) Placement of the newly added Gene Ontology term GO:0097447 'dendritic tree', drawn using the ontology editing tool OBO-Edit (see ‘Methods’). Due to space limitations, not all ancestor and descendant terms are shown. Is_a links are indicated by "I"; part_of links are indicated by "P" (see main text for explanation). Has_part links are also discussed in the main text.
The newly added GO-CCO terms (integrated from SAO) include cytoplasmic inclusions such as ‘Lewy body’ and subtypes, cell-type specific variants of structures such as ‘CA3 hippocampus pyramidal cell dendrite’ and terminal boutons such as ‘C bouton’ (Figure 3). Very specific terms such as ‘CA3 hippocampus pyramidal cell dendrite’ have been included because they represent instances with peculiarities that influence their biological role, e.g. because they convey particular electrical properties. The full list of newly added terms is available in Table 1.

Representation of 'synapse part' in the Gene Ontology. Children of the GO term. GO:0044456 'synapse part' are shown using the ontology editing tool OBO-Edit (see ‘Methods’). Terms underlined in green were added as part of the SAO amalgamation. Due to space limitations, not all ancestor and descendant terms are shown. Is_a links are indicated by "I" (see main text for explanation).
Some SAO terms represented multicellular anatomical terms, and were therefore out of scope for the GO. Instead, they were suggested as additions to ontologies such as Uberon (e.g. ‘axon bundle’, defined as ‘Group of myelinated or unmyelinated axons that group together.’).
Some recent extensions that make use of additional spatial information
Has_part
In addition to is_a and part_of within the GO-CCO, the GO also makes use of additional relationship types, both within the GO-CCO and connecting the GO-CCO to other ontologies.
Recently the has_part (BFO:0000051) relation was added to the GO-CCO [17] to represent the composition of components with respect to their sub-components. These relationships add value beyond the reciprocal part_of relationships because they add a dependency of a structure to always have a certain sub-part. While it is true to say that every nucleus is part of a cell at every point during the existence of that nucleus, it is not true that every cell has a nucleus (exceptions including bacterial cells and mammalian erythrocytes) – thus the GO-CCO includes a part_of link between nucleus and cell, but no reciprocal has_part link. Similarly, there is a has_part link between ‘trans splicesomal complex’ and ‘U2 snRNP’, but no reciprocal part_of link. U2 snRNPs are not always a part of a trans splicesomal complex, but every trans splicesomal complex has a U2 snPNP part.
Connecting cell components to cell types
To record the connections between terms in the GO-CCO and cell types in the cell type ontology, the GO maintains an additional supplementary bridging ontology called x-cell-parts.owl[18] that contains statements such as ‘astrocyte projection’ part_of some ‘astrocyte’. The former term belongs to the GO and the latter to the Cell Ontology (CL) [10, 19]. In addition, the Cell Ontology includes links from cell types such as ‘nucleate erythrocyte’ to GO-CCO terms such as ‘nucleus’. These can be found in the full version of the Cell Ontology [20].
Connecting biological processes and molecular functions to cell components
Logical definitions (equivalence axioms) are being added to the GO that link the Biological Process (BP) and Molecular Function (MF) branches of the ontology to the GO-CCO [21]. These additional axioms are available in an extended version of the GO called go-plus [22], and allow for fuller reasoning over GO as well as for automation of new term creation using the GO TermGenie system [manuscript in preparation]. This work is ongoing, and axioms are being added using relationships such as results_in_assembly_of, results_in_disassembly_of, occurs_in, has_start_location, capable_of. Some examples of such relationships in OWL are shown below. For illustrative purposes we use a variant of OWL Manchester Syntax where we include labels in with the identifiers to enhance readability.
Class: ‘GO:0000045 ! autophagic vacuole assembly’ EquivalentTo:‘GO:0022607 ! cellular component assembly’ and results_in_assembly_of some ‘GO:0005776 ! autophagic vacuole’
Class: ‘GO:0000390 ! spliceosomal complex disassembly’ EquivalentTo: ‘GO:0022411 ! cellular component disassembly’ and results_in_disassembly_of some ‘GO:0005681 ! spliceosomal complex’
Class: ‘GO:0006264 ! mitochondrial DNA replication’ EquivalentTo: ‘GO:0006260 ! DNA replication’ and ‘BFO:0000062 ! occurs_in’ some ‘GO:0005739 ! mitochondrion’
Class: ‘GO:0006888 ! ER to Golgi vesicle-mediated transport’ EquivalentTo: ‘GO:0016192 ! vesicle-mediated transport’ and has_target_end_location some ‘GO:0005794 ! Golgi apparatus’ and has_target_start_location some ‘GO:0005783 ! endoplasmic reticulum’
Terms that describe protein complexes and integration with the Protein Ontology
Of the current 3332 terms in the GO-CCO, 1622 terms are descendants of GO:0032991 macromolecular complex; most of these (1510) are descendants of GO:0043234 protein complex.
Like the rest of GO, protein complexes in the GO-CCO should be applicable to more than one species. This can be difficult to achieve when a complex has been characterized in a single species, or homology among species is unclear. In these cases our aim is to make the term as generic as possible. A protein complex from a particular species is often described in the GO-CCO textual definition, but the scope of that term is not limited to that species. Other resources can use these generic protein-complex terms to classify more specific entities. For example, the Protein Ontology (PRO) [23] makes species-specific protein complex subclasses of the generic GO protein complexes.
An example of a protein complex, the mouse-specific form of eukaryotic translation initiation factor 2 complex 1, is represented in PRO using the ID PR:000026828 (Figure 4). Each species-specific translation initiation factor complex is composed of specific protein entities, which can be any combination of isoforms, processed forms, or post-translationally modified forms. In this example, the mouse complex is shown. The human complex would be defined using human proteins and would be restricted to the human taxon. Both human and mouse complexes are defined with an is_a relationship to the generic GO complex. At present, there are almost 500 protein complex terms in PRO [24].

Protein ontology report for entry PR:000026828. Ontology information for the Protein Ontology term 'eukaryotic translation initiation factor 2 complex 1 (mouse)', showing parent GO term and has_part links to other PR terms [25].
GO-CCO protein complexes are defined by some combination of their biological function, their subunit composition in one or more species, and their location within the cell. Protein complexes in the GO range from simple dimeric complexes, for example ‘TAP complex’, to complexes having many subunits, for example ‘proteasome complex’. Homodimeric complexes are also included.
In the protein complex branch of the GO-CCO, most protein complexes (729 of 1502) are direct subclasses of ‘protein complex’ itself. This ‘flat’ arrangement is not ideal for the purposes of navigation and data summarization. GO curators are working with the IntAct group [26] to improve the sub-categorization of protein complexes. In cases where a protein complex always contributes to a larger macromolecular structure in a cell we provide a part_of relationship between the protein complex and the larger component. For example, ‘histone deacetylase complex’ is part of the nucleoplasm in the GO. In cases where complexes are found in multiple locations, or move between cell components, the generic protein complex is merely part of ‘cell’.
Applications of the CC ontology
GO Annotation
The primary use of the GO-CCO is to annotate localization of gene products. There are currently 886238 annotations (both experimental and electronic) of gene products from a variety of species. These annotations can be interrogated with GO browsers such as AmiGO 2 [27, 28] or QuickGO [29, 30]. Annotation describes the process of assigning GO terms to gene products. Annotation can be carried out either automatically or manually. Automated methods provide a fast and efficient way of creating a large set of annotations. For automatic annotation, curators have constructed various mapping files between external features and GO terms. GO annotations are automatically applied to gene products via the mapping files. For example, InterPro entries are manually annotated with terms from the GO [31]. InterPro entry IPR019038 (DNA polymerase subunit Cdc27) contains a mapping to GO:0005634 ‘nucleus’, and any protein that is a member of this InterPro family will receive the annotation GO:0005634. Similarly, UniProtKB entries are manually and electronically tagged with keywords [32] including a cellular component category. A mapping file between UniProtKB keywords and GO terms allows transitive electronic GO annotation of the UniProtKB entry.
Manual annotations are created by curators assessing experimental evidence from published literature. Manual annotations generally result in the use of more specific GO terms. Curators use the experiments or analyses described in a paper to infer the localization of a gene product in a particular cellular compartment. For example, a curator has annotated the human MPV17 mitochondrial membrane protein-like protein (MPV17L; UniProtKB:Q2QL34) with the GO term GO:0005777 'peroxisome' using evidence from Iida et al., [33] who demonstrate co-localization of MPV17L with a fluorescent peroxisomal marker.
Sequence-based analysis can also form the basis for an annotation; GO-CCO annotations can be made based on the presence of a key sequence or structural feature in the gene product. Manual annotations can also be transferred to similar gene products either computationally or when orthology is indicated in the literature. Whether automatic or manual, every annotation is attributed to a source (either a literature reference, a computational analysis or another database) and an evidence code is used to indicate the type of evidence that supports the annotation [34].
Recently, the GO annotation model has been extended to include contextual annotations. A GO annotation can be further refined using ontology terms from within the GO or other ontologies. For example, the PomBase annotation for the ‘cut8’ gene to ‘proteasome localization’ (a GO biological process term) is further refined by specifying that this localization takes place in the ‘nuclear membrane’ (a GO-CCO term) (Figure 5).

Refining GO annotations using Gene Ontology CC terms. The PomBase annotation for the ‘cut8’ gene to ‘proteasome localization’ (a GO biological process term) is further refined by specifying that this localization takes place in the ‘nuclear membrane’ (a GO-CCO term). (Taken from http://www.pombase.org/spombe/result/SPAC17C9.13c#go).
Neuronal connectivity
Neuron connectivity maps provide a way to help understand how the nervous system works. The FlyBase anatomy ontology contains a variety of connectivity relationships between neurons in the Drosophila nervous system [35] and these relationships are used to deliver powerful queries using the Virtual Fly Brain database [36]. These relationships use the GO-CCO to precisely specify how the cells are connected – for example, the has-pre-synaptic-terminal-in relation is specified using the GO-CCO class ‘post-synaptic membrane’.
Image annotation
The Cell Image Library (CIL) is a database of cell images that is indexed using multiple ontologies [6]. The GO-CCO is the ontology that is used to annotate any subcellular structures highlighted in the image. The CIL can be searched using GO terms, and the hierarchy of the GO is incorporated into the query. For example, searching for ‘cytoskeleton’ (GO:0005856) returns images annotated directly with ‘cytoskeleton’, as well as cytoskeletal parts, such as ‘microtubule basal body’ e.g. http://www.cellimagelibrary.org/images/38899.
Pathway and interaction databases
The GO-CCO has been used in a variety of pathway databases such as IntAct and Reactome to specify the site in which binding or a molecular event takes place.
IntAct [26], a member of the International Molecular Exchange Consortium (IMEX), uses the GO-CCO to capture molecular interaction data. The IntAct database [37] can record the site of interaction by cross-referencing interaction entries to GO-CCO terms. Over 8000 experimentally-defined IntAct interactions have manually-assigned GO-CCO terms and users can search on interaction cross-references to retrieve, for example, all interactions that occur at the plasma membrane (GO:0005886) or nucleus (GO:0005634).
Because protein function and location are often tightly linked, the manually curated and peer-reviewed pathway database Reactome [38] cross-references GO-CCO terms [39]. Reactome uses a subset of the GO-CCO to annotate the subcellular locations of entities; entities include proteins, nucleic acids, small molecules and subatomic particles, and can be a single molecule or a collection of components such as a macromolecular complex. A GO-CCO term is also a required attribute in the curation of a Reactome event, a biochemical reaction with a defined input (reactant) and output (product), such as the association of two proteins to form a complex, or a catalytic reaction.
Annotation of models
The EBI BioModels database [40, 41] contains a number of systems biology models represented in SBML (Systems Biology Markup Language) format. SBML provides slots for indicating the compartment of a reaction, resulting in the ability to search for models involving particular cellular components, such as the ‘giant axon’.
Enhanced search of multiple resources
The GO-CCO can also be used to search a variety of databases via the Neuinfo interface [16]. For example, searching for data associated with “axoneme” (GO:0005930) [42] returns several data types, including images of axonemes and aging phenotypes associated with axonemes.
Phenotypes and disease
Although the GO-CCO, like the rest of the GO, focuses on structures that are found mostly in ‘non-pathological’ contexts, the GO-CCO has applications in bioinformatics analyses of phenotypes and diseases. The Mammalian Phenotype (MP) ontology [43] contains terms for describing abnormalities observed in clinical or model organism experimental settings. Many MP terms such as ‘abnormal mitochondrion morphology’ have been translated to OWL expressions that reference GO-CCO terms such as ‘mitochondrion’, allowing for cross-database phenotype comparisons [44]. The Neurodegenerative Disease Phenotype Ontology (NDPO) [7], with its associated Phenotype Knowledge Base (PKB), uses a model that incorporates descriptions for both human disease phenotypes and those of animal models. Entities are drawn from community ontologies (including the GO-CCO) made available through the Neuroscience Information Framework (NIF) and qualities are drawn from the Phenotype and Trait Ontology (PATO). The resulting phenotype statements describe structural alterations at the subcellular, cellular and gross anatomical levels.
Many diseases are the result of an abnormality within a specific cell component. For example, the disease class ‘ciliopathy’ encompasses a range of disorders such as Bardet-Biedl syndrome and Alström syndrome characterized or caused by an abnormality of the cilium or its subcellular structures. Candidate genes for disorders such as these can be found by scanning GO annotations for gene products that are localized to, or play some role in, the assembly of the relevant cellular components.
Currently the GO-CCO has relationships to taxa of the form ‘only in taxon’ or ‘never in taxon’ [45, 46]. For example, the CC ‘plastid’ has a never_in_taxon link to ‘Metazoa’ and ‘Fungi’ supported by a particular publication (in this case [47]). We intend to increase the coverage of GO for certain kinds of unicellular organisms that are currently under-represented in GO. For example, trophozoites such as Giardia have characteristic structures such as a 'ventral disc', which can be further subdivided into other parts such as microribbons and microtubular components. These additions will be accompanied by the relevant taxon constraints [45]. Our priority is to include new terms as required for annotation. Other possible areas for extension include dinoflagellates and algae.
Conclusions
For researchers to be best able to make use of the data available to them, a single system for classification is essential. Prior to this work, the NIF’s SAO and the GO-CCO provided alternative systems for classifying the same data, potentially hindering cross-database analyses. The SAO has now been incorporated into the GO-CCO, which was expanded where necessary, with the result being a single system of classification for subcellular entities across both resources. The SAO was never intended, however, to replicate the GO; rather, it was a means of specifying the relationships among structures encountered in microscopic images. As such, it was a model for describing instances [3, 48], rather than intending to serve as a reference ontology. With the improvement in tools for working with community ontologies, in particular, tools to convert between OBO format and OWL (http://oboformat.org) it no longer made sense to maintain the two separately. Rather, the SAO will be rebuilt as an annotation model that imports the GO-CCO for cellular components.
The enhancements to the Cellular Component section of the GO described in this paper will benefit researchers in basic biology, biomedicine and systems biology who use ontologies in their research. The amalgamation of the SAO into the GO-CCO resulted in a single unified ontology designed to serve the broad needs of GO annotators as well as the specific needs of the neuroscience community.
Methods
Ontology Development
The SAO was originally developed using Protégé 3, and the GO-CCO developed as part of GO using OBO-Edit [49]. More recently the GO editors have been using a hybrid approach, developing GO using a combination of OBO-Edit, Protégé 4 and TermGenie [manuscript in preparation]. In particular, a supplementary constraints ontology called x-disjoints.owl is maintained in OWL and used as part of the GO continuous integration system [50] using OWL reasoners such as Elk [51]. This ontology contains OWL axioms stating, for example, that a nucleus and a cytoplasm can share no parts.
Amalgamation
In order to amalgamate the SAO into the GO-CCO, we supplemented the manually maintained mappings between the SAO and the GO-CCO (which had become stale since the SAO was first developed) with automatically-generated mappings based on lexical matching. These were all individually examined by GO editors to determine their accuracy. If considered correct, they were added into the GO-CCO with a database cross-reference to SAO.
For the remaining SAO terms for which no mapping to the GO-CCO could be determined manually or automatically, the GO editors evaluated each to determine if it was appropriate to add to the GO. See the section ‘Amalgamation with SAO’ for more details.
Availability
The URL for the Gene Ontology (GO) is http://www.geneontology.org/.
Gene Ontology files are publicly available for download [52]. Detailed documentation on the Gene Ontology can also be found online [53]. Researchers wishing to annotate their experiments using GO terms may refer to the GO’s annotation guidelines [54], and contact the GO helpdesk [55].
Abbreviations
- BP:
-
Biological process
- CC:
-
Cellular component
- CCDB:
-
Cell centered database
- GO-CCO:
-
Gene ontology cellular component ontology
- CIL:
-
Cell image Library
- CL:
-
Cell ontology
- GO:
-
Gene ontology
- MF:
-
Molecular function
- MP:
-
Mammalian phenotype
- NDPO:
-
Neurodegenerative disease phenotype Ontology
- NIFSTD:
-
Neuroscience information framework standard
- OBO:
-
Open biological ontologies
- OWL:
-
Web ontology language
- SAO:
-
Subcellular anatomy ontology.
References
The Gene Ontology.http://www.geneontology.org/,
Ashburner M, Ball CA, Blake JA, Botstein D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT, Harris MA, Hill DP, Issel-Tarver L, Kasarskis A, Lewis S, Matese JC, Richardson JE, Ringwald M, Rubin GM, Sherlock G: Gene ontology: tool for the unification of biology. The Gene Ontology Consortium. Nat Genet. 2000, 25: 25-29. 10.1038/75556.
Larson SD, Fong LL, Gupta A, Condit C, Bug WJ, Martone ME: A formal ontology of subcellular neuroanatomy. Front Neuroinformatics. 2007, 1: 3-
Gupta A, Bug W, Marenco L, Qian X, Condit C, Rangarajan A, Müller HM, Miller PL, Sanders B, Grethe JS, Astakhov V, Shepherd G, Sternberg PW, Martone ME: Federated access to heterogeneous information resources in the Neuroscience Information Framework (NIF). Neuroinformatics. 2008, 6: 205-217. 10.1007/s12021-008-9033-y.
Foundational Model of Anatomy ontology - Home.http://sig.biostr.washington.edu/projects/fm/index.html,
Martone ME, Tran J, Wong WW, Sargis J, Fong L, Larson S, Lamont SP, Gupta A, Ellisman MH: The cell centered database project: an update on building community resources for managing and sharing 3D imaging data. J Struct Biol. 2008, 161: 220-231. 10.1016/j.jsb.2007.10.003.
Maynard SM, Mungall CJ, Lewis SE, Imam FT, Martone ME: A knowledge based approach to matching human neurodegenerative disease and animal models. Front Neuroinformatics. 2013, 7: 7-
Imam FT, Larson SD, Bandrowski A, Grethe JS, Gupta A, Martone ME:Development and use of ontologies inside the neuroscience information framework: a practical approach.Front Genet. 2012, 3: 111-
Smith B, Ceusters W, Klagges B, Köhler J, Kumar A, Lomax J, Mungall C, Neuhaus F, Rector AL, Rosse C: Relations in biomedical ontologies. Genome Biol. 2005, 6: R46-10.1186/gb-2005-6-5-r46.
Bard J, Rhee SY, Ashburner M: An ontology for cell types. Genome Biol. 2005, 6: R21-10.1186/gb-2005-6-2-r21.
Cooper L, Walls RL, Elser J, Gandolfo MA, Stevenson DW, Smith B, Preece J, Athreya B, Mungall CJ, Rensing S, Hiss M, Lang D, Reski R, Berardini TZ, Li D, Huala E, Schaeffer M, Menda N, Arnaud E, Shrestha R, Yamazaki Y, Jaiswal P: The plant ontology as a tool for comparative plant anatomy and genomic analyses. Plant Cell Physiol. 2013, 54: e1-10.1093/pcp/pcs163.
Zebrafish Anatomy Ontology.http://zfin.org/action/ontology/show-all-anatomy-terms,
Hayamizu TF, Mangan M, Corradi JP, Kadin JA, Ringwald M: The adult mouse anatomical dictionary: a tool for annotating and integrating data. Genome Biol. 2005, 6: R29-10.1186/gb-2005-6-3-r29.
Mungall CJ, Torniai C, Gkoutos GV, Lewis SE, Haendel MA: Uberon, an integrative multi-species anatomy ontology. Genome Biol. 2012, 13: R5-10.1186/gb-2012-13-1-r5.
Bug WJ, Ascoli GA, Grethe JS, Gupta A, Fennema-Notestine C, Laird AR, Larson SD, Rubin D, Shepherd GM, Turner JA, Martone ME: The NIFSTD and BIRNLex vocabularies: building comprehensive ontologies for neuroscience. Neuroinformatics. 2008, 6: 175-194. 10.1007/s12021-008-9032-z.
Neuroscience Information Framework.http://neuinfo.org/,
Gene Ontology Consortium: The Gene Ontology in 2010: extensions and refinements. Nucleic Acids Res. 2010, 38: D331-D335.
GO-Cell Ontology cross-products file.http://purl.obolibrary.org/obo/go/extensions/x-cell-parts.owl,
Meehan TF, Masci AM, Abdulla A, Cowell LG, Blake JA, Mungall CJ, Diehl AD: Logical development of the cell ontology. BMC Bioinformatics. 2011, 12: 6-10.1186/1471-2105-12-6.
Cell Ontology OWL.http://purl.obolibrary.org/obo/cl.owl,
Mungall CJ, Bada M, Berardini TZ, Deegan J, Ireland A, Harris MA, Hill DP, Lomax J: Cross-product extensions of the gene ontology. J Biomed Inform. 2011, 44 (1): 80-86. 10.1016/j.jbi.2010.02.002.
GO extended version, go-plus.owl.http://purl.obolibrary.org/obo/go/extensions/go-plus.owl,
Natale DA, Arighi CN, Barker WC, Blake JA, Bult CJ, Caudy M, Drabkin HJ, D’Eustachio P, Evsikov AV, Huang H, Nchoutmboube J, Roberts NV, Smith B, Zhang J, Wu CH: The protein ontology: a structured representation of protein forms and complexes. Nucleic Acids Res. 2011, 39: D539-D545. 10.1093/nar/gkq907.
Bult CJ, Drabkin HJ, Evsikov A, Natale D, Arighi C, Roberts N, Ruttenberg A, D’Eustachio P, Smith B, Blake JA, Wu C: The representation of protein complexes in the Protein Ontology (PRO). BMC Bioinformatics. 2011, 12: 371-10.1186/1471-2105-12-371.
PR:000026828 report.http://purl.obolibrary.org/obo/PR_000026828,
Kerrien S, Aranda B, Breuza L, Bridge A, Broackes-Carter F, Chen C, Duesbury M, Dumousseau M, Feuermann M, Hinz U, Jandrasits C, Jimenez RC, Khadake J, Mahadevan U, Masson P, Pedruzzi I, Pfeiffenberger E, Porras P, Raghunath A, Roechert B, Orchard S, Hermjakob H: The IntAct molecular interaction database in 2012. Nucleic Acids Res. 2012, 40: D841-D846. 10.1093/nar/gkr1088.
Carbon S, Ireland A, Mungall CJ, Shu S, Marshall B, Lewis S, AmiGO H: Web presence working group: AmiGO: online access to ontology and annotation data. Bioinforma Oxf Eng. 2009, 25: 288-289.
Binns D, Dimmer E, Huntley R, Barrell D, O’Donovan C, Apweiler R: QuickGO: a web-based tool for Gene Ontology searching. Bioinforma Oxf Engl. 2009, 25: 3045-3046. 10.1093/bioinformatics/btp536.
QuickGO.http://www.ebi.ac.uk/QuickGO/,
Burge S, Kelly E, Lonsdale D, Mutowo-Muellenet P, McAnulla C, Mitchell A, Sangrador-Vegas A, Yong S-Y, Mulder N, Hunter S: Manual GO annotation of predictive protein signatures: the InterPro approach to GO curation. Database J Biol Databases Curation. 2012, 2012: bar068-
UniProtKB Keywords.http://www.uniprot.org/keywords/,
Iida R, Yasuda T, Tsubota E, Takatsuka H, Matsuki T, Kishi K: Human Mpv17-like protein is localized in peroxisomes and regulates expression of antioxidant enzymes. Biochem Biophys Res Commun. 2006, 344: 948-954. 10.1016/j.bbrc.2006.04.008.
Guide to GO Evidence Codes.http://www.geneontology.org/GO.evidence.shtml,
Osumi-Sutherland D, Reeve S, Mungall CJ, Neuhaus F, Ruttenberg A, Jefferis GSXE, Armstrong JD: A strategy for building neuroanatomy ontologies. Bioinforma Oxf Engl. 2012, 28: 1262-1269. 10.1093/bioinformatics/bts113.
Milyaev N, Osumi-Sutherland D, Reeve S, Burton N, Baldock RA, Armstrong JD: The Virtual Fly Brain browser and query interface. Bioinforma Oxf Engl. 2012, 28: 411-415. 10.1093/bioinformatics/btr677.
IntAct Database.http://www.ebi.ac.uk/intact/,
Reactome.http://www.reactome.org/ReactomeGWT/entrypoint.html,
Matthews L, Gopinath G, Gillespie M, Caudy M, Croft D, de Bono B, Garapati P, Hemish J, Hermjakob H, Jassal B, Kanapin A, Lewis S, Mahajan S, May B, Schmidt E, Vastrik I, Wu G, Birney E, Stein L, D’Eustachio P: Reactome knowledgebase of human biological pathways and processes. Nucleic Acids Res. 2009, 37: D619-D622. 10.1093/nar/gkn863.
Li C, Donizelli M, Rodriguez N, Dharuri H, Endler L, Chelliah V, Li L, He E, Henry A, Stefan MI, Snoep JL, Hucka M, Le Novère N, Laibe C: BioModels Database: An enhanced, curated and annotated resource for published quantitative kinetic models. BMC Syst Biol. 2010, 4: 92-10.1186/1752-0509-4-92.
BioModels Database.http://www.ebi.ac.uk/biomodels-main/,
NIF - axoneme Search Results.https://neuinfo.org/mynif/search.php?q=axoneme,
Smith CL, Eppig JT: The mammalian phenotype ontology as a unifying standard for experimental and high-throughput phenotyping data. Mamm Genome Off J Int Mamm Genome Soc. 2012, 23: 653-668. 10.1007/s00335-012-9421-3.
Mungall CJ, Gkoutos GV, Smith CL, Haendel MA, Lewis SE, Ashburner M: Integrating phenotype ontologies across multiple species. Genome Biol. 2010, 11: R2-10.1186/gb-2010-11-1-r2.
Clark JI D n, Dimmer EC, Mungall CJ: Formalization of taxon-based constraints to detect inconsistencies in annotation and ontology development. BMC Bioinformatics. 2010, 11: 530-10.1186/1471-2105-11-530.
Kuśnierczyk W: Taxonomy-based partitioning of the gene ontology. J Biomed Inform. 2008, 41: 282-292. 10.1016/j.jbi.2007.07.007.
Miyagishima S: Mechanism of plastid division: from a bacterium to an organelle. Plant Physiol. 2011, 155: 1533-1544. 10.1104/pp.110.170688.
Chen L, Martone M, Gupta A, Fong L: OntoQuest: exploring ontological data made easy. Proc. 32nd Int. Conf. Very Large Data Bases. 2006, Seoul, South Korea, 1183-1186.http://www.vldb.org/2011/,
Day-Richter J, Harris MA, Haendel M, Lewis S: OBO-Edit–an ontology editor for biologists. Bioinforma Oxf Engl. 2007, 23: 2198-2200. 10.1093/bioinformatics/btm112.
Continuous Integration of Open Biological Ontology Libraries | Bio-Ontologies SIG.http://bio-ontologies.knowledgeblog.org/405,
elk-reasoner - A Java-based OWL 2 EL reasoner.http://code.google.com/p/elk-reasoner/,
GO Ontology Downloads.http://geneontology.org/GO.downloads.ontology.shtml,
Gene Ontology Documentation.http://geneontology.org/GO.contents.doc.shtml,
GO Annotation: Policies and Guidelines.http://www.geneontology.org/GO.annotation.shtml,
Contact GO.http://www.geneontology.org/GO.contacts.shtml,
Acknowledgements
This work was supported by the National Human Genome Research Institute (NHGRI) P41 grant 5P41HG002273-09 to the Gene Ontology Consortium and by the European Union RTD Programme ‘Quality of Life and Management of Living Resources’ QLRI-CT-2001-00981 and QLRI-CT-2001-00015 to GO and UniProtKB-GOA groups at EMBL-EBI. HJD is supported by HG002273, and National Institute of General Medical Sciences (NIGMS) GM080646). JL is funded by the European Molecular Biology Laboratory (EMBL), European Bioinformatics Institute Outstation (EMBL-EBI) core funds. Support for the Neuroscience Information Framework is provided by a contract from the NIH Neuroscience Blueprint HHSN271200800035C via the National Institute on Drug Abuse. CJM's contribution was supported by the Director, Office of Science, Office of Basic Energy Sciences, of the U.S. Department of Energy under Contract No. DE-AC02-05CH11231.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that no competing interest exists.
Authors’ contributions
PR worked on the SAO-GO-CCO amalgamation and drafted the manuscript. DPH and JL contributed to the SAO-GO-CCO amalgamation. MEM and FTI worked on the NIF project, on the SAO and on the CCDB. HD is also a curator for the Protein Ontology and contributed to the section on protein complexes. PR, DPH, TZB, REF, HD, CJM and JL worked on expanding the GO-CCO and linking it to other branches of GO and to external ontologies. MEM, DPH, TZB, REF, HD, CJM and JL helped draft the manuscript. All authors read and approved the final manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under license to BioMed Central Ltd. This is an open access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Roncaglia, P., Martone, M.E., Hill, D.P. et al. The Gene Ontology (GO) Cellular Component Ontology: integration with SAO (Subcellular Anatomy Ontology) and other recent developments. J Biomed Semant 4, 20 (2013). https://doi.org/10.1186/2041-1480-4-20
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/2041-1480-4-20